

Note : Le présent chapitre n’a pas vocation à faire du lecteur un expert en génétique, pour cela je conseille le livre « Hérédité chez l’abeille et les colonies d’abeille » de Bernard Sauvager (Editions ANERCEA).
Chez l’abeille, à l’instar de la plupart des êtres vivants le support de l’information génétique est réparti sur deux sites : l’ADN (acide désoxyribonucléique) nucléaire présent dans le noyau de toutes les cellules (qui ont un noyau) qui composent l’organisme et l’ADN mitochondrial (Figure 1). Ces deux marqueurs différents mais complémentaires permettent d’identifier la diversité génétique de l’abeille.

L’ADN mitochondrial est un petit peu moins connu et requiert quelques explications. La mitochondrie est l’équivalent de la centrale énergétique de la cellule. Ce composant cellulaire (organite) n’est transmis que par les mères. Ainsi l’ADN de votre mitochondrie est celui de la mère de la mère … de votre mère. Appliqué aux abeilles, l’ADN mitochondrial permet donc de suivre les filiations maternelles de la reine. Le mâle ne transmet que la moitié des gènes de l’ADN nucléaire.
Ainsi, dans l’ADN nucléaire en revanche, il y aura un mélange de gènes du faux bourdon et de la reine. Une reine étant polyfécondée, un brassage génétique très important peut être observé à ce niveau entre la
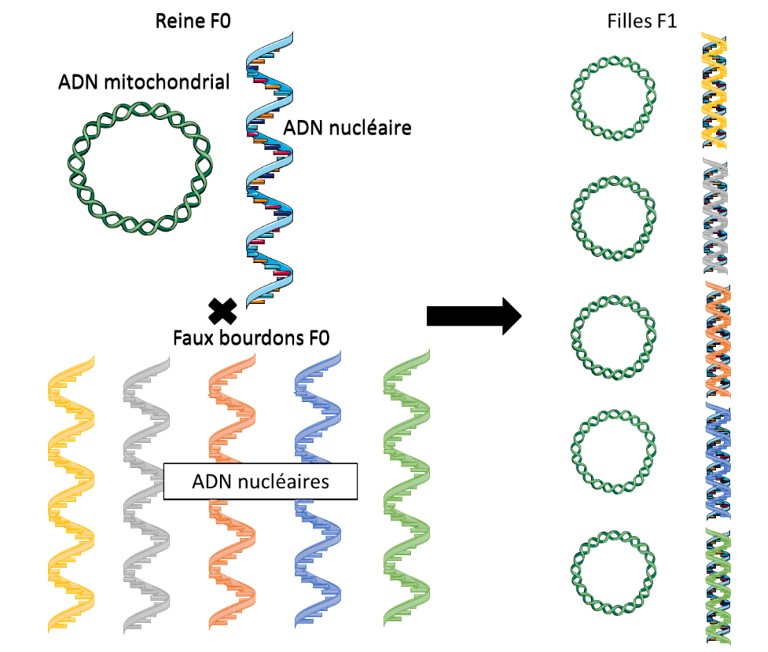
descendance fille (F1) au sein de la colonie (Figure 2). Ce marqueur permet donc d’identifier le niveau d’introgression (d’hybridation) des lignées.
On déduit de la Figure 2 que :
- Un seul spécimen suffit à déterminer la génétique mitochondriale de la colonie (la reine a transmis à sa descendance son génome mitochondrial)
- De multiples specimens seront nécessaires à l’analyse de l’introgression du génome nucléaire. Pour cette étude, il est habituel d’analyser des séquences d’ADN très spécifiques appelées microsatellites.
Historiquement, Apis mellifera a initialement été classée en plusieurs lignées sur la base des caractères morphologiques[1] :
- A : lignée Africaine
- M : lignée Ouest et Nord européenne (l’abeille « noire »)
- C : lignée Sud et Est européenne
- O : lignée Orientale (Caucase, Turquie, Moyen Orient, Chypre et Crète)
La génétique mitochondriale a permis de confirmer cette classification tout en introduisant, actuellement, 2 nouvelles lignées :
- Y : Ethiopie[2]
- S ou Z : Proche Orient (Syrie)[3]
La répartition géographique globale et française de ces lignées est décrite en Figure 3 gauche et droite respectivement.
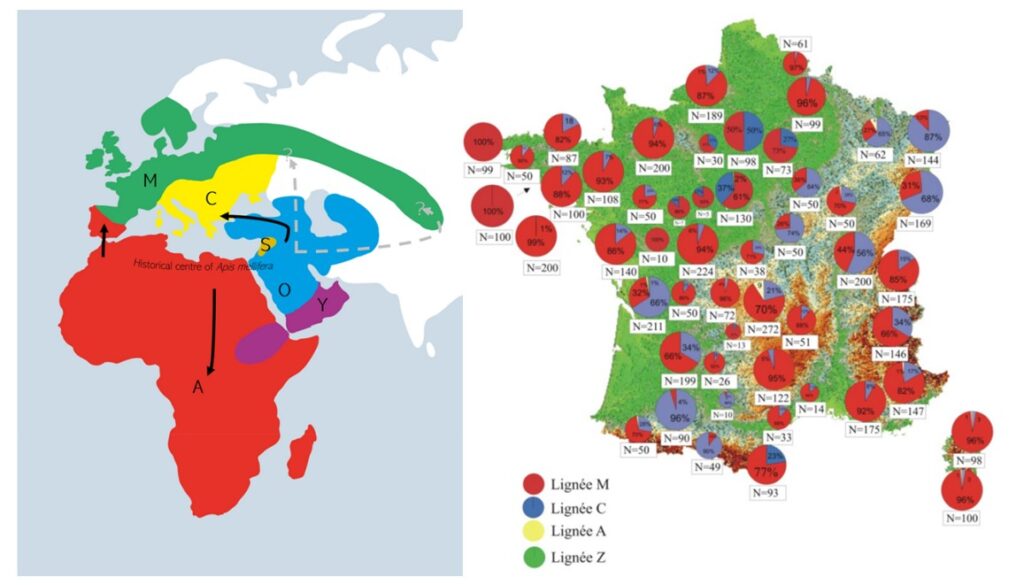
Figure 3: Répartition globale (gauche) et française (droite) des lignées d’Apis mellifera. La figure de gauche est issue des travaux de Tihelka, Cai [4] et du Bilan du cheptel Français de l’abeille domestique (2004- 2006)
L’ADN mitochondrial ne connait pas de brassage génétique et n’évolue donc principalement que par des mutations. Le levier de sélection génétique est donc d’utiliser la plasticité du code génétique nucléaire, et pour cela nous avons besoin de « bons » mâles. Génétiquement les mâles descendent d’un œuf non fécondé de la reine… ou d’ouvrières dans le cas d’une ruche dite bourdonneuse. Non fécondé signifie qu’ils ne possèdent que la moitié du code génétique soit 16 chromosomes, ils sont alors qualifiés d’haploïdes (les femelles : ouvrières et reine sont diploïdes et ont 2x 16 = 32 chromosomes). Au niveau de la transmission génétique cela entraîne des conséquences lourdes (Figure 4).
Toutes les filles d’un mâle auront donc la moitié de leur génome nucléaire qui sera identique à celui de leur géniteur mais seulement 50% de celui de leur mère. Ainsi les sœurs auront au moins 50% de leurs gènes identique (apport du père), alors que les demi-sœurs peuvent avoir des gènes complétement différents (gènes paternels différents et gènes maternels aléatoires).
En résumé : une sélection sur la reine permet de gérer la lignée mitochondriale héréditaire A, C, M, O mais ne permet pas de prédire les caractéristiques de la descendance. Une « bonne » reine pourra générer de reines intéressantes pour la sélection uniquement si les mâles apportent les allèles attendus. C’est aussi pour cela que le greffage peut donner, du fait de la paternité multiple, des reines au comportement très différent.
Bibliographie :
- Ruttner, F., Apis mellifera Linnaeus 1758:576 General Introduction to a Polymorphic Species with an Unusual Range of Adaptation, in Biogeography and Taxonomy of Honeybees, F. Ruttner, Editor. 1988, Springer Berlin Heidelberg: Berlin, Heidelberg. p. 165-175.
- Franck, P., et al., Genetic diversity of the honeybee in Africa: microsatellite and mitochondrial data. Heredity, 2001. 86(4): p. 420-430.
- Alburaki, M., et al., Mitochondrial structure of Eastern honeybee populations from Syria, Lebanon and Iraq. Apidologie, 2011. 42(5): p. 628-641.
- Tihelka, E., et al., Mitochondrial genomes illuminate the evolutionary history of the Western honey bee (Apis mellifera). Scientific Reports, 2020. 10(1): p. 14515.